
Breeding & Genetics
Part Two - Color Mutations in Parrots
Note: this article focuses primarily on psittacines (parrots). Some of the information is applicable to other bird and animal species.
Article sections:
PART ONE - NORMAL COLORATION
Basic elements of bird coloration
1. Feather layers
2. Melanin
3. Red/yellow pigments
a. UV markings and fluorescence
4. Structural color
5. Pigment in the eyes
a. Strange eye colors in lutino cockatiels
b. Unexplained red eyes
Color production and deposition
1. Melanin production
2. Carotenoid pigments
3. Psittacin production
4. Structural color
PART TWO - COLOR MUTATIONS IN PARROTS
Melanin-altering
mutations Psittacin-altering mutations Distribution-altering
mutations Structure-altering
mutations Non-inheritable genetic variations Combination
mutations
A note on leucism, albinism,
and dilution
1. Sex-linked lutino
a. Parino mutations
(lutino alleles) - lime/pallid/platinum
2. Cinnamon
3.
Fallow mutations (including recessive silver and NSL ino)
4.
Dilute mutations (including emerald)
5. Dominant edged (including dominant silver)
6. Faded/Isabel
7. Melanistic
1. Blue (whiteface)
a. Parblue mutations
2. Orangeface/tangerine/dominant yellow cheek
3. Sex-linked yellow
cheek and goldcheek
4. Red-fronted and
red suffused
1. Pied
a. Dominant
b. Recessive (ADM)
c. Progressive
d. The pied mutation and eye color
2. Opaline/pearl
a. Is it ancestral
coloring?
b. Digression - are green
cockatoos possible?
1. Dark factor/olive
2. Grey/greygreen
3. Violet
4. Other structural mutations
1. Grizzle
2. Halfsiders and tricolor
1.
Albino (blue lutino), creamino (parblue lutino)
2. Lacewing (cinnamon lutino)
3. Rubino (opaline lutino)
4. Cinnamon fallow
All life forms on earth have a lot of the same genes, and the more closely related one life form is to another, the more similar their genes will be. It's estimated that humans have 7% of the same genes as bacteria, 15% the same as mustard grass, 85% the same as zebra fish, and 98% the same as chimpanzees (The Tech).
The 350+ species in the parrot family are more closely related to each other than they are to any other bird group or other life form, and naturally they share a lot of the same genes. We don't know how many genes are involved in color production but the number must be dozens if not hundreds.
Some of these genes are obviously different from each other, since every parrot species looks at least somewhat different than any other. But all parrot species use the same basic elements to produce color (melanin, psittacin, and feather structure) and the same basic process to produce these elements, generally controlled by the same genes. A specific mutation in one of these common genes will have the same effect in all species that share the gene. The visual result might not always be the same depending on what the bird's normal color looks like, but it will have the same effect on the underlying color production process. Most species don't have every single mutation that is currently known, but they have the potential to develop these mutations.
So far there has not been a comprehensive sequencing of the parrot family genome, which would tell us precisely what genes the different species have in common. This could lead to an identification of the physical location of mutation genes, and enable us to confirm whether similar-looking mutations in different species are caused by the same gene or not. We can hope that this will happen in the not too distant future, but until then we have to rely on observation, logic, and a certain amount of guesswork.
Color mutations in parrots can be broken down into four basic categories:
- Mutations that change the quality or quantity of melanin pigment
- Mutations that change the quality or quantity of psittacin pigment
- Mutations that change the distribution pattern for pigment.
- Mutations that change the feather structure without affecting the quality or quantity of pigment in the feather
Normal bird coloration is the subject of active scientific study, and the books Bird Coloration Volumes 1 & 2 by Hill and McGraw are a major source of information on the state of our knowledge as of 2006. But so far the subject of mutation coloring in birds has not attracted the attention of the "official" scientific community. There are some studies of mutation coloring in other animals, mostly mammals, but their applicability to birds is questionable. Most of our information on parrot mutations comes from the work of well-informed volunteer hobbyists, primarily Terry Martin, an Australian veterinarian who has written two books on the subject, and Mutavi, a research group centered in the Netherlands that performs electron-microscope examinations of feathers and other tissues to obtain information about mutation effects. It was Mutavi who made the determination that parrots have only eumelanin pigment and not pheomelanin; they do not have the capability to perform the chemical tests that would confirm this, but pheomelanin has a different shape than eumelanin and they feel confident that they could detect the difference with the electron microscope. A complete list of the members of Mutavi is not publicly available, but Inte Onsman and Dirk van den Abeele have been the most active in writing articles for public viewing.
Specific mutations and their effect on the color production process will be discussed in the following sections. Major sources of information include Martin's parrot mutations book and cockatiel mutations book, Mutavi, and Martin's now-defunct Genetics-Psittacine Yahoo group, whose old discussions were available online until Yahoo cut back on group functions in 2020. Even though this information has now disappeared, I have kept the dead links in the article as a reminder of where I got the information.
*******************
(Back to Top)
We don't have a standardized set of terminology to describe parrot mutations. A mutation that's called by one name when it occurs in one species might be called by a different name in a different species, and by yet another name in a third species. Conversely, the name that's used for a specific mutation in one species might be used to describe a different mutation in a different species. There are a lot of mutations in a lot of different species, so these terminology differences can be very confusing. If you're used to the mutation terminology for one species and get involved in a conversation about a different species, it can almost seem like a different language sometimes.
The conflicting terminology is partly due to the fact that it's often hard to recognize that a mutation that has a different visual effect in different species is actually the same mutation, and the mutation names were chosen long before anyone knew about the connection. Some of the mutation names are "green-centric", meaning that they describe what happens when a certain mutation occurs in a parrot that is normally green, and it doesn't make sense to apply the name to a parrot that is NOT normally green.
A prime example of this is the blue mutation, which removes the psittacin (red/yellow pigment). If this species normally has green feathers, removing the yellow pigment will leave you with blue feathers; hence the name. But removing the yellow pigment from a cockatiel won't result in a bird with blue feathers; instead you get a grey bird whose face is white instead of yellow and orange. A blue and gold macaw is normally blue to begin with, and in this species the blue mutation results in a bird that is still the same shade of blue but has lost the gold coloring. A parblue (partial blue) mutation called seagreen removes some of the psittacin. In a normally green parrot it subtracts some yellow pigment, resulting in a bird that is greenish blue with more green than blue. In a crimson rosella, the subtracted pigment is red not yellow, and the visual result is that the bird looks orange instead of red. But the technically correct name for this mutation is still seagreen, even though the bird doesn't have any green feathers. You can see why the name "blue mutation" didn't catch on in species where the coloring doesn't change to blue.
There is a movement to standardize the terminology, spearheaded by Terry Martin and his associates. But it hasn't made a lot of headway. There is an international agreement on mutation naming posted on Genetics-Psittacine with an updated version posted on Ornitho-genetics. But the international group that made this agreement is small and there aren't very many people who know about the agreement. A lot of the terminology requires a fairly advanced knowledge of coloration mechanics and mutation genetics to be understandable. It's green-centric too so it's not a natural way to talk about non-green species. Much of the old terminology has been in use for decades and is firmly entrenched with the general public, who know little and care less about the finer points of mutation terminology, so the old mutation names will probably continue to prevail for a long time.
For easy understandability, this article will primarily use the common North American names for mutations, making note of widely used alternate names. Most of my birds are cockatiels, so there's a reason for it if my descriptions seem cockatiel-centric - I'm just writing what I know. I'm also catering to my audience, since a lot of my internet activity is on cockatiel groups.
*******************
(Back to Top)
These mutations interfere with the melanin-producing process in some way, changing the physical properties of the pigment or the way it is deposited. For information on the normal color production process, please see Part One.
A note on leucism, albinism and dilution. Leucism is sort of a catch-all term for reduced pigment and different sources define it in different ways. Some sources say leucism affects all pigments and others (including Terry Martin's book on parrot mutations) say it specifically affects melanin, and other pigments are disregarded. There are similar multiple definitions for albinism.
Albinism is usually defined as the more or less total absence of melanin (or pigment in general). Leucism can be less extreme, and may remove just some of the pigment so there's still a color tint to the feathers. There are different types of leucism that have different effects, so there don't seem to be any hard and fast rules about when a bird with reduced pigment can be called leucistic and when it can't. Many of the mutations described in the following sections could be described as a type of leucism, and so could a number of individual aberrations that are not a recognized mutation.
BTO says that carotenoid coloring in birds isn't affected by leucism. But carotenoid pigments are obtained through the diet so it's a different situation than birds who make their yellow/red pigments internally.
Terry Martin presented this definition in an article:
"Albinistic mutations correlate with mutant alleles of loci involved in the process of melanin production including metabolism and the construction of melanosomes. The hallmark of an albinistic mutation, compared to the other two groups, is the reduction in melanin within soft tissue and not just feathers. In other words, they lose melanin from eyes, skin, legs, beak and toenails. Some of these colour morphs have dark eyes when mature, but all show signs of reduced eye colour upon hatching. Melanin granules produced by these colour morphs are altered in structure, the change varies depending upon the function of the locus involved, but all show qualitative melanin changes.
"Dilution mutations correlate with mutant alleles of loci controlling deposition of melanin from the melanocytes into the melanosomes of the feather. They are able to produce normal melanin granules, but do not transfer normal quantities into feathers. They are said to have quantitative changes to their melanin levels. Soft tissues are unaffected because the melanin deposition process in the eyes, skin and other tissue is different to the feathers. Put more simply, the dilute loci have no function in skin melanin deposition.
"Leucistic mutations correlate to mutant alleles of loci that control melanocyte life functions, in particular migration of melanoblasts (melanocyte precursors) from the neural crest in the embryo. True leucistic mutations are uncommon, the black-eyed clear phenotype being the true appearance. But partial leucism is very common. These are Pied birds; those that have some regions devoid of melanin and yet other regions fully coloured. The hallmark of leucistic colour morphs is that melanin is either totally absent or fully retained. There are never qualitative nor quantitative changes to melanin in these colour morphs. Many mutant alleles with leucistic characteristics also show evidence of pleiotrophic behaviour. This means they have effects beyond the basic effect on melanin production and is not surprising for loci controlling complex functions like melanoblast migration."
In summary, there are basic three ways we can alter the melanin content of a feather. Damage the cells responsible for its production resulting in leucism, damage the metabolic process of melanin production resulting in albinism, or damage the transfer process for melanin granules from the melanocyte into target areas in the feather resulting in dilution.
1. Sex-linked lutino
Alternate common names: ino
Internationally agreed name: lutino, although it was felt that ino green
(for a yellow lutino) or ino blue (for an albino) is technically more correct. Some sources prefer yellow series ino
or white series ino to avoid green-centricity. No official word on the
preferred terminology for a bird that primarily uses red psittacin rather
than yellow, but the word rubino is commonly used to describe these birds.
Inheritance mode: sex-linked recessive
Sex-linked lutino is one of the most common and widespread mutations in parrots. There is one other lutino mutation called non-sex-linked (NSL) lutino but it is much less common. Because it is the predominant lutino mutation, sex-linked lutino is usually referred to simply as lutino without specifying that it is sex-linked, while NSL ino always specifies that it is NSL. Sex-linked lutino makes melanin coloring disappear from view, usually completely, although a melanin color wash may appear on the feathers when lutino is combined with certain other mutations (especially cinnamon). Any structural coloring stops functioning due to the absence of normal melanin, so that only the psittacin coloring remains visible. A lutino bird looks yellow in species that primarily use yellow psittacin, and the word lutino loosely translates as "yellow one".
The sex-linked ino locus codes for MATP, which is a membrane transporter protein whose job it is to transport tyrosinase, TRP1 and TRP2 into the melanosome. In the lutino mutation the MATP protein is defective and unable to function properly. It is able to transport small amounts of these three substances, so some melanin is produced and deposited in the feather. But there is not enough of it to be visible to the human eye. The protein matrix that supports the pigment is severely deformed. Tyrosinase activity is 2.5 times higher than normal. (Genetics-Psittacine, Lovebirds Compendium page 387). Mutavi has an electron microscope picture of the deformed melanin in a lutino budgie feather.
In a different article, Mutavi reports that the protein matrix is composed of a number of different proteins, each one controlled by a different gene. A mutation in any one of these genes can result in the normal "product" not being delivered, with the result that the matrix will be deformed and too small. This in turn affects the amount of pigment that can be deposited on the matrix. The linked article does not specifically discuss the lutino mutation, but it looks like this principle is applicable.
a. Parino mutations (lutino alleles) - Lime/Pallid/Platinum
The parino (partial ino) mutations are alleles of the lutino locus
(different variations of the lutino gene). They basically do the same
thing that the lutino gene does, but they don't do it to such an extreme
degree, so the visual effect is different. The
Allelic Mutations article
has a more detailed explanation.
The number of parino mutations is not
entirely clear, but there are at least two. Lime/pallid produces darker coloring than platinum, which
produces darker coloring than lutino, which usually produces no visible
melanin coloring.
Lovebirds Compendium page 420 says that pallid reduces the melanin by
about 50% and platinum reduces it by 75%. It is not always clear which specific allele the alternate names below
refer to; however the platinum mutation in cockatiels is apparently the same as
the "official" platinum mutation.
Ornitho-genetics indicates that there is a third allele called pearly,
and a mutation called pale is also
listed as a possible allele of SL ino.
Lovebirds Compendium page 420 says that pearly reduces the melanin by
about 90% and pale reduces it by about 15%.
Alternate common names: clearbody (in budgerigars); Isabel (scarlet-chested parrots, peachfaced
lovebirds); also Australian ino or Australian cinnamon in peachfaced
lovebirds in some locations; lacewing (Indian ringecks); yellow (swift
parrot); dark-eyed cinnamon (quaker parrot); fallow (green-cheeked conure);
pallid (in Europe). The red-rumped parrot has two variations; the darker
version is called lime and the paler version is called platinum.
Internationally agreed name: pallid (for the
darker version) and platinum (for the paler version).
Inheritance mode:
Sex-linked. Recessive to the wildtype (normal) allele, co-dominant with
the mutation alleles including lutino.
As alleles of SL lutino, these mutations would also involve a disruption in
the coding for the MATP protein, but the disruption is less severe and
enough melanin is deposited in the feather to produce visible melanin
coloration or structural color involving melanin. The melanin-based color is paler than normal. In a green
bird, this means that the blue coloring is less intense while the yellow
remains the same, and the combined effect is a more yellowish shade of green
than normal.
2. Cinnamon
Alternate common names: none,
apparently, although there are several unrelated mutations that are incorrectly
being called cinnamon.
Internationally agreed name: cinnamon
Inheritance mode: sex-linked recessive
This is another widespread, common mutation. Melanin that would normally be black has a brownish color instead, although in some species (like cockatiels) the visual effect often looks more like a paler shade of grey instead of brown to us. This might be because the basic feather structure influences the appearance of colors. An alternative explanation is that eumelanin that has not been converted all the way to black is often described as a grey-brown color. We usually describe the color as brown, but it might look more like grey in some species, mutations, or individuals.
The cinnamon locus codes for TRP1, an enzyme which acts during the final stage of eumelanin production to change the color of the pigment from brown to black. In the cinnamon mutation the bird cannot produce this enzyme, so this stage can not be completed and the finished eumelanin looks brown instead of black. Cinnamon does not 'produce' brown, it prevents the normal development of black (Genetics-Psittacine).
*******************
(Back to Top)
3. Fallow mutations (including recessive silver and NSL ino)
There are several different fallow mutations that have a fairly similar effect on coloration, but they are not caused by the same gene and have different mechanisms behind their visual effect. The word fallow essentially describes the phenotype (physical appearance) not the genotype (genes). Fallow is defined as a mutation that is autosomal recessive in inheritance; causes red eyes in adult birds; and changes all black pigments to light grey and/or brown shades without affecting the physical properties of any other pigment or the physical distribution of pigment. Some fallow mutations eliminate the white iris ring found in some species.
A more general definition is that fallow is a type of albinism in which there is a generalized decrease in the amount of melanin in the eyes, feathers and skin. All areas have a normal number of melanocytes but the ability of those cells to produce melanin is either reduced (in fallows) or absent (in NSL ino) (Mutavi).
The loose definition of fallow and the difficulty in determining which specific gene was responsible for the color in a given species has resulted in the word being used pretty loosely in naming mutations.
Terry Martin's cockatiel mutations book reports that fallow-type morphs are produced in humans by the tyrosinase locus; the adapter protein 3 (AP-3) locus, the P protein locus, and the membrane-associated transporter protein (MATP) locus. The tyrosinase locus is involved in bronze fallow/NSL ino, but it is not clear which if any of these other loci are involved in the other fallow mutations.
Terry Martin's parrot mutations book lists four kinds of fallow:
- Bronze fallow (simply called fallow in the US without the word
"bronze"), which reduces black pigment to brown. NSL ino is an
allele of this mutation.
Ornitho-genetics
indicates that there are two other alleles of this gene called dark-eyed clear and
pastel. These mutations reduce the level of tyrosinase activity, and
it's reduced to zero in the case of NSL ino. The protein matrixes
are normal, but in NSL ino the
matrix remains empty and uncolored, with no
pigment attached (Lovebirds Compendium
page 345). Some pigment is present in the other alleles but it may be clumped,
irregular and of poor quality (Mutavi).
In bronze fallow, the decrease in melanin granules is 50% in the barbules
and 75% in the barbs; in pastel the decrease is 50% in the barbules and about
100% in the barbs (B-F).
The location of this gene is called the a-locus.
NSL ino is common in some parrot species, but it may have ceased to exist in cockatiels. It was never common in this species, and the cockatiel community has apparently lost track of the last known breeder (Cockatiel Chromatics). Sometimes the attempt to establish a new mutation fails.
- Pale fallow, which reduces black pigment to light brown (as described by Martin). However "pale fallow" is used in Europe to describe cockatiels that Terry Martin classifies as ashen fallow, and it's believed that pale fallow and ashen fallow might actually be the same mutation. Pale fallow affects the protein matrix in melanin granules, possibly due to the lack of one of the necessary proteins. The matrix is small and deformed, which affects the amount of pigment that can be deposited on it (B-F, Mutavi). Tyrosinase activity is higher than normal. The location of this gene is called the pf-locus.
- Ashen fallow (called recessive silver in the US and
pale fallow in Europe), which reduces black pigment to light grey
(as described by Martin).
The locus name is not reported on
Ornitho-genetics and page 472 of
Lovebirds Compendium
(authored by the owner of the Ornitho-genetics website) says
"As far as I know, this 'ashen or smokey fallow' is only present in cockatiels... I personally have my doubts about the existence of this 'ashen fallow' as a separate mutation because the cockatiel [Nymphicus hollandicus] has a different feather structure than most other parakeets... to just compare these 'visually' with birds with feathers of the structural type does not seem to be ideal. But anyway, to be absolutely certain of whether this 'ashen fallow' exists or not a lot of research needs to be conducted."
The book indicates that the color distinctions for all the fallow mutations were determined visually, which is subjective and prone to error. The cinnamon mutation (which results in melanin that is brown not black) tends to look like light grey rather than brown in cockatiel feathers, so it wouldn't be surprising if pale fallow (which results in light brown melanin) tended to look like light grey in cockatiels instead of showing its true color.
- Dun fallow, which reduces black pigment to grey-brown. The location of this gene is called the df-locus. We don't know how this mutation affects the melanin production process. In lovebirds this mutation produces a limited melanin reduction in the feathers, and an almost complete reduction of melanin in the eyes (Lovebirds Compendium page 493).
An
alternative explanation is that eumelanin that has not been converted
all the way to black is often described as a grey-brown color. We
usually call it brown, but it might look more like grey in some species,
mutations, or individuals.
Terry Martin also wrote an article on fallow morphs which can be viewed on Archive.org when the website is feeling cooperative.
Because these different fallow mutations are caused by different recessive genes, breeding a bird with one fallow mutation to a bird with a different fallow mutation will produce offspring who are normal with splits, not visual fallow.
*******************
(Back to Top)
4. Dilute mutations (including emerald and marbled)
"Dilute" is another category of melanin-altering mutations which are grouped together based on their effect on physical appearance not on the gene or the mechanism involved in creating this appearance. Dilute mutations are autosomal recessive mutations that greatly reduce the amount of melanin pigment in the feathers, but the eyes always remain dark. In at least some species, the beak, legs and feet have the normal amount of melanin too; the source materials don't indicate whether this is a universal characteristic. Some species have multiple dilute mutations that belong to the same genetic family, while other dilute mutations are unrelated. Common names for dilute mutations include yellow, pastel, suffused, and silver (Martin). The greywing/clearwing/dilute mutations in budgies are alleles of the same locus.
Dilute mutations affect the quantity of melanin pigment not its physical properties. In general it would appear that there is a problem with transporting the completed melanin granule into the feather. Possible reasons include abnormalities in the dendrites or in the transport protein myosin.
The emerald mutation in cockatiels is a dilute mutation which has been shown to have defective dendrites, and a similar pattern to the greywing and clearwing mutations in budgies. The melanin granules can't leave the melanocyte normally due to the defective dendrites so they build up, causing the melanocyte to swell and eventually rupture, spilling melanin granules into the skin. The result is an irregular rate of pigment deposit into the growing feather, with large globs taken up at times and very little at other times. (R. Solis on American Cockatiel Society).
The Australia-only pastel silver cockatiel mutation is also a dilute mutation.
There is a recessive edged dilute mutation that is best known in lovebirds but occurs in several other species including the Australia-only silver spangle cockatiel. The official internationally-agreed name for this mutation has been changed to marbled for all species (Ornitho-Genetics, GenCalc).
5. Dominant edged (including dominant silver)
Alternate common names: Edged dilute, dominant dilute. Dominant silver is
often called zoomed or gezoomed by European cockatiel breeders.
Internationally agreed name: dominant edged
Inheritance mode: Dominant or co-dominant to the normal (wildtype) gene;
in some species the
effect is more pronounced with double factor birds than with single factor.
Dominant edged is generally considered to be a diluting mutation, meaning that it reduces the quantity
of melanin without affecting its physical properties; but it is not
considered to be a dilute mutation of the type described in the previous
section. The inheritance mode is different and the way it affects the amount
of pigment is different.
The quantity of pigment in the feather is variable, so that the color is typically darker near the edges of the flight feathers where the melanin is more dense, and paler in the middle of the flight feathers, creating the appearance of a dark border. There is also some degree of "scalloping" on the body feathers, but some body parts may have a darker color overall than others. In the dominant silver cockatiel, DS juveniles can be difficult to distinguish from normal grey, but the markings become more noticeable over time. The body coloring becomes more diluted while the back of the head often remains dark, creating a "skullcap" effect. Cocks lighten up more than hens do.
The mechanism behind this progressive color change is unknown but early melanocyte death is one hypothesis. Another hypothesis involves the PMEL protein, a matrix protein that is transported by MATP (Genetics-Psittacine-1 Genetics-Psittacine-2), in which case this would really be a type of albinism that affects melanin quality, rather than a diluting mutation. The dendrites are normal, unlike the emerald mutation (described in the previous section) which produces coloration so similar to DS that sometimes people ask whether dominant silver and emerald are the same mutation. (Martin-parrots, Martin-cockatiels, R. Solis on American Cockatiel Society)
Yu et al report that during feather growth, melanin is deposited first near the edge of the feather and then the melanocytes gradually withdraw to "feed" the more centrally located cells. They also note that the barbules near the edge of the feather are usually more heavily pigmented with melanin than those in the center. The dominant edged coloring may be a manifestation of this pattern that is more exaggerated than normal.
*******************
(Back to Top)
"Faded" seems to be another general class of mutations which have a similar effect, rather than a specific mutation involving the same gene in all species. According to Martin-Parrots, faded mutations are recessive in inheritance. They slightly reduce the amount of black pigment, often resulting in a brownish, cinnamon-like look. But in reality the melanin in the feather is still black. These mutations are often called Isabel (which has no real definition and is often applied to mutations that don't qualify as faded). "Recessive cinnamon" is frequently used too, but this is not a true cinnamon mutation since the melanin color is normal. Although a number of species have faded mutations, it does not seem to be particularly widespread.
*******************
(Back to Top)
This is yet another general category of mutations that have a similar effect but are probably caused by different genes in at least some cases. Melanistic mutations are uncommon and have only been observed in a handful of species. These mutations cause melanin pigment to spread into areas where it is not normally found. According to Terry Martin, it's likely that these mutations are caused by the malfunction of a gene that normally restricts melanin distribution and holds it to a specific pattern. If the increased melanin is deposited in the core of the feather, it can result in structural colors like green, blue and purple appearing in areas that are normally yellow, white, or red (if the feather structure needed to produce these colors is present). The blue-fronted mutation in the rainbow lorikeet is an example of this. If the increased melanin is deposited on the outer layer of the feather, there will be black coloring in areas where it is normally not found. The melanistic Eastern Rosella and the blackface budgerigar are examples of this.
*******************
(Back to Top)
We can talk about the visual effect of psittacin-altering mutations but not much else, since we know nothing about the way psittacin is produced and very little about its chemical properties. So we can't tell exactly what these mutations do to disrupt the normal process. The blue mutation and the related parblue mutations seem to be the only common psittacin-altering mutations, although there are a few other psittacin-altering mutations that are only known to affect a few species. This suggests that psittacin production might be simpler than melanin production, with fewer genes involved.
According to Terry Martin on Genetics-Psittacine,
1. Blue
Alternate common names: whiteface (in cockatiels)
Internationally agreed name: blue
Inheritance mode:
autosomal (ordinary) recessive
The blue mutation does not create blue color; rather it removes the yellow and red pigment, usually throughout the body but sometimes only in some areas. In a parrot that was mostly green to begin with, removing the yellow will result in a blue bird. In a parrot that was NOT mostly green to begin with, the action of the gene will not produce feathers that look blue. In these species, removing the yellow will result in the confusion that comes from having a mutation name (blue) that does not actually match the color of the bird. In cockatiels this problem was solved by calling the mutation whiteface instead of blue, since that describes the visual result of removing the yellow and orange color from the bird's face. But the location of this gene on its chromosome is called the blue locus in all species. The assumption seems to be that if you know enough about genetics to be talking about a locus, you're knowledgeable enough to understand that a blue-locus mutation doesn't necessarily result in a blue bird.
In budgies there are two different blue mutations which both appear to be mutations of the blue locus. They are considered to be heteroalleles of this locus, meaning different variations of the same gene that can occur in the same gene pair, but they do not interact with each other the same way that alleles of the same locus usually do. One of these mutations is the common, widespread "standard" blue mutation that removes all psittacin from the body, while the second one is apparently known only in budgies at present and leaves some yellow psittacin in the face. The subject is beyond the scope of this article, but Bergman discusses it in detail. Please see the Allelic Mutations article for an explanation of alleles.
Recent research by Cooke et al has identified the location of the blue locus. It's on Chromosome 1, within an 11-gene region bounded by SNPs at positions 21,019187 and 21,445,705. The gene in this region that's most likely responsible is called LOC101880715, which they have nicknamed MuPKS (for Melopsittacus undulatus PolyKetide Synthase). You have no idea how happy it makes me to have trivia like this. This is the first time that the precise location of a parrot mutation gene has been identified.
They also identified the specific DNA change that makes the psittacin disappear. There's a substitution of a single amino acid at a specific location. The blue birds have only tryptophan at this location, and the green birds have arginine at this location (either homozygous for arginine or heterozygous for both arginine and tryptophan). The researchers were able to produce yellow psittacofulvin in yeast using cloned budgie cells.
They proposed a biochemical mechanism for yellow psittacofulvin
pigment synthesis in budgerigars, including a proposed explanation for how
the blue mutation disrupts it. It's too technical for most of us:
"The ACP is activated by attachment of phosphopantetheine. Chain initiation involves transacylation of an acetyl-primer unit from acetyl-CoA to the active-site cysteine of the ketoacyl synthase (KS) domain. Each cycle of chain elongation begins with a condensation reaction between the KS-bound growing polyketide chain and malonyl-ACP. The MAT domain is responsible for transferring malonyl extender units from malonyl-CoA to the ACP. The ketoreductase (KR) and dehydratase (DH) domains convert each β-ketothioester intermediate to the corresponding u nsaturated α,β-unsaturated thioester. The inactive ER domain of MuPKS cannot reduce this double bond, resulting in a conjugated polyene product, such as those observed in the feather pigments of the scarlet macaw."
a. Parblue mutations (blue alleles) - Aqua/Turquoise
In addition to the two alleles that are considered to be "true" blue, there
are at least two parblue (partial blue) mutations which remove some but not
all of the psittacin pigment. These parblue mutations are recessive to the
wildtype gene and co-dominant to each other (Genetics-Psittacine),
and to the "standard" blue mutation; they are homoalleles that do
interact with each other as expected (Bergman,
Allelic Mutations article).
As mentioned earlier, alleles are different variations of the same gene, and the parblue mutations are different variations of the blue gene. The parblue mutations do the same thing that the blue gene does but they don't do it to such an extreme degree, so the visual effect is different. They reduce the psittacin coloring but don't eliminate it. In a bird that is normally green, the parblue mutations result in blue-green coloring, with the exact shade depending on how much the psittacin was reduced.
The aqua mutation reduces the psittacin pigment by about 50%. It appears that aqua is the same as the pastelface mutation in cockatiels. The turquoise mutation reduces the psittacin to about 20% and is equivalent to creamface in cockatiels (Cockatiel Chromatics). There are various other names used for the parblue mutations and their combinations. Seagreen seems to be the preferred term when a bird has one gene for aqua and one for turquoise.
*******************
(Back to Top)
2.
Orangeface/tangerine/dominant yellowcheek
Alternate common names: dominant yellowcheek (in cockatiels)
Internationally agreed name: orangeface
Inheritance mode: co-dominant to the normal gene, but many
authorities prefer to classify it as recessive in peach-faced lovebirds
because the single-factor coloring is difficult to detect in birds with
other mutations. In cockatiels
it is accepted as a dominant or co-dominant mutation
Although Terry Martin prefers to call this the tangerine locus, I'm not aware of any species where it is called the tangerine mutation. This mutation apparently affects red psittacin but not yellow. In lovebirds, the face color is changed from rosy to orange. The color of the tail markings changes too, while the body color is thought to remain unchanged (although some sources report a subtle change in body color). In lovebirds, a double factor bird (one that has two copies of the gene) has a different appearance than single factor (a bird with only one copy of the gene). In cockatiels, the cheek color is changed from orange to yellow or yellow-orange, although the other splits and mutations that the bird has will have an influence on the appearance of the cheek spot. There may be individual variations as well that are not related to the presence of other mutations. Because of this variability in appearance, it's difficult to tell exactly how different a DF cockatiel is from an SF cockatiel. Terry Martin's cockatiel book (published in 2007 and possibly outdated on this subject) says that adult cocks change their cheek color (becoming more orange), and that double factor birds may be more likely to have a purer yellow face, with DF cocks perhaps being more likely to retain the yellow color in adulthood.
Because of all the questions about inheritance mode and the action of the gene, and because it's only been observed in two species, I wonder if orangeface and DYC are really mutations of the same locus. It might be the action of species-specific genes that coincidentally have a similar effect, in a couple of species whose red psittacin occurs solely or primarily in the face.
*******************
(Back to Top)
3.
Sex-linked yellowcheek and goldcheek
Alternate common names: yellowface (for SLYC)
Internationally agreed name: yellowcheek (for SLYC)
Inheritance mode: sex-linked
recessive for SLYC; sex-linked dominant for goldcheek
Sex-linked yellowcheek is only known to occur in cockatiels and may involve a gene that's unique to this species, involved in producing the orange cheek spot. This mutation reportedly produces a cheek spot color that is a more pure yellow than in the dominant yellowcheek mutation. But once again the other mutations and splits that the bird has will affect the cheek color, and sometimes it can be difficult to tell the difference between DYC, SLYC, and pastelface. In particular, a split to whiteface tends to produce a more pronounced orange color in the yellowcheek mutations.
It is reported that SLYC hens are poor mothers, to the point that foster parents are often required to successfully raise the offspring of an SLYC hen. It is suspected that this mutation might affect the hormone balance somehow, particularly the estrogen level.
The goldcheek mutation may be another psittacin-altering mutation that is unique to cockatiels. This mutation has always been very rare and not much is known about it. It's said that these birds are difficult to breed, and the hens are bad mothers who refuse to sit. Some people consider this mutation to be functionally extinct although there are still some surviving individuals. The visual appearance is somewhat similar to the yellowcheek mutations but the cheek color has more of a dull golden tone. It was originally thought to be a parblue mutation, but test breeding results indicate that it is actually sex-linked dominant (R. Solis in American Cockatiel Society newsletter for Oct-Dec 2014, Vol. 38 No. 4).
It is possible that SLYC and goldcheek could be alleles (different variations of the same gene). They have a lot in common - both are sex-linked and the hens are poor mothers in both cases. One is recessive to the normal and the other is dominant, but I see no reason why this couldn't happen with alleles. But we will probably never know the answer, because it isn't practical to test breed these two mutations with each other.
*******************
(Back to Top)
4. Red-fronted and red-suffused
Alternate common names: red-bellied (for red-fronted)
Internationally agreed name: ???
Inheritance mode: There's no agreement. It's recessive
in the turquoise
parrot according to
Wikipedia.
Martin-Parrots
indicates that this mutation is dominant to the normal yellow belly coloring
in the turquoise parrot and scarlet-chested parrot, saying that it's
possible to breed yellow-bellied birds from two red-bellied birds, but you
can not breed a red-bellied bird from two yellow-bellied birds.
GenCalc says it's co-dominant in princess parrots,
and incomplete dominant in the
scarlet-chested parrot and
turquoise parrot.
These mutations increase the amount of red psittacin in a bird and allow it to appear in places where it's not normally found. Red-fronted primarily increases the red or orange coloring on the chest and belly, while red suffused has red/orange coloring scattered throughout the plumage or as barring. There is little information on these mutations so it's not clear how many different genes may be involved. The amount of red coloring varies considerably from one individual to another, and it's suspected that there are a number of modifier genes that influence the expression of the primary gene. Parent birds with more red tend to produce offspring with more red. Abnormal red coloring can also be caused by health and diet issues, so it's not always easy to distinguish between genetic red coloring and that caused by external factors.
Species that may have a true red-fronted or red-suffused mutation include the turquoise parrot, scarlet-chested parrot, swift parrot, and princess parrot (Martin-Parrots).
*******************
(Back to Top)
When parrot mutations are categorized, pied and opaline (pearl) are usually described as mutations that alter the distribution and/or pattern of pigment. This seems a bit arbitrary to me, since many of the mutations categorized as melanin-altering or psittacin-altering affect the distribution pigment of in some way, and some of these mutations (like dominant edged) create a pattern on the feather. But sticking with familiar established tradition is easier on everyone than getting into a pointless discussion about how these mutations should really be classified. In any case, the way pied changes the distribution of melanin is radically different than the way the melanin-altering mutations do it, so it's reasonable to classify it differently. The case for opaline is less clear-cut.
Pied and opaline both reduce the amount of melanin in the feathers, but it has been observed that the amount of psittacin increases at the same time. As far as I know, this does not occur with any of the mutations classified as melanin-altering. It is hypothesized that melanin inhibits the expression of psittacin somehow (possibly a paracrine effect), and when the melanin itself is inhibited in certain ways the psittacin is able to express itself more strongly (GeneticsPsittacine-1, GeneticsPsittacine-2).
It appears to me that just the attempt to produce melanin is enough to trigger this inhibiting effect, since we do not see an increase in psittacin in a lutino. With the lutino mutation there is an attempt to produce melanin, but even though it fails pretty miserably it is evidently enough to keep the psittacin in check. The primary feathers of cockatiels provide a nice demonstration of this effect. In normal hens and juveniles, the grey primaries have yellow spots on them. On a lutino the spots are still visible, but they are a more intense yellow than the rest of the feather; apparently the psittacin is inhibited in the area where there was an attempt to produce and deposit melanin, but it is not inhibited in the small areas that are melanin free. There was no attempt to produce melanin in a pied feather, and a pied primary cockatiel feather has no markings; the color is uniform throughout. The normal markings are visible in lutino cockatiel tail feathers, and pearl markings are visible on a lutino pearl bird. None of these markings appear on a pied feather. But on a lutino pied cockatiel, the pied feathers can be a noticeably brighter shade of yellow than the non-pied feathers.
In some species the increase in psittacin coloring is dramatic. In the Bourke's parrot, the opaline mutation changes a mostly-brown bird (melanin coloring) into a mostly-pink bird (psittacin coloring) - the rosy Bourke. In the Congo African Grey, a certain type of pied mutation results in large patches of bright red feathers in areas that are normally grey without any red at all - the red factor CAG.
*******************
(Back to Top)
There are a number of different pied mutations caused by a variety of different genes. They have one thing in common: they affect the differentiation and migration of melanoblasts in the embryo, so that some areas of the skin lack melanoblasts (and ultimately melanocytes). The melanin-producing cells are absent so there is no melanin-producing capability in these areas. But these mutations affect the melanoblasts in different ways. Lovebirds Compendium page 499 reports that at least 15 different pied mutations have been identified in mice. Some affect the production of pigment cells, while others might influence the lifespan of the melanoblasts, or affect the skin tissues surrounding the pigment cells so that they can no longer function.
The information on the mechanics of pied coloration are rather sketchy for parrots. But this condition is called piebaldism in mammals and other non-avian species, and Genetics Home Reference says that it can be caused by mutations in the KIT and SNAI2 genes, which are genes that birds possess. It can be caused by a variety of other genes and conditions as well, but it appears to me that these are the genes that are primarily involved in healthy piebaldism not associated with genetic disease. The KIT gene in particular looks like a good candidate for the cause of at least some of the pied mutations in birds (GHR). However Parichy et al question the role of KIT in the timing of avian melanoblast migration, and they list several other factors that could have an effect.
One previous hypothesis about piebald coloring was that it is caused by directed migration of melanoblasts, which traveled slower than usual and failed to arrive in some areas. But Mort et al found that this was not the case in a study on mice. Instead they reported that migration was undirected and migration speed was normal with the piebald mutation, but a KIT mutation caused reduced proliferation of the melanoblasts. Some areas lack pigmentation because not enough melanoblasts developed within the necessary time frame to populate the area.
The melanoblasts involved in feather/skin coloration originate in the neural crest (the precursor to the spinal cord) at shoulder level and migrate into the rest of the body following the dorsolateral (back and sides) path (Parichy et al). The melanoblasts that have to travel the furthest tend to be affected most by the pied mutation. Pied feathers occur most frequently at the wingtips (primary feathers) and lower body (Genetics-Psittacine). Conversely, in a heavily pied cockatiel the area that is most likely to retain melanin coloring is the "saddle" area of the upper wings, which is closest to the origin point of the melanoblasts.
Martin-Parrots identifies four different pied mutations: dominant, recessive, recessive anti-dimorphic (ADM), and recessive black-eyed clear. Several years later on Genetics-Psittacine Martin said "Since that time it has become quite apparent that all recessive Pied mutations in parrots have ADM features and I now believe they all represent a single locus which I would refer to as Recessive Pied." Therefore recessive pied and recessive ADM pied will be treated as a single mutation in this article.
Some pied mutations can reduce psittacin in some species. Therefore it appears that some pied genes may work on loci that are pleiotropic (have more than one function). This is not well understood. Pied may affect the iris color in some species.
*******************
(Back to Top)
a.
Dominant pied
There are at least two forms of dominant pied, both of which occur in
budgies and are called Dutch pied and Australian (banded) pied according to Martin-Parrots. However
the Dutch pied seems to be little known, and budgie mutation websites
generally ignore it and instead talk about another type of dominant pied,
clearflight (Cute
Little Birdies,
Budgieplace).
Al-Nasser
provides more information on the history of Dutch pied and clearflight,
indicating that Dutch pied is only common in Australia. The links in this
paragraph do a good job of explaining the differences between these
mutations and they also do a good job of explaining dark-eyed clear,
so I won't try to rehash the information.
Recessive pied markings do not follow a standard pattern and
the markings can be highly variable although as explained earlier, certain
areas of the body tend to be affected more than others in accordance with
the migration path followed by melanoblasts in the embryo.
At present there are no species known to have more than one type of
recessive pied, so it's not known whether all forms of recessive pied are
caused by the same gene or if there are multiple recessive pied mutations. In addition to the "major"
recessive pied gene,
Terry Martin believes that there are 10-15 pied "modifier" genes, some dominant and
some recessive, that influence the expression of the major recessive pied
gene but do not express themselves in the absence of the major gene.
When a single copy of the major gene is present, the minor genes may express
themselves in small ways, such as the "tickmark" on the back of the head
that is common in split-pied cockatiels (ANCS).
Another possible explanation for the visible signs of a split is that there
is some degree of co-dominance or incomplete penetrance.
Recessive pied disrupts the development of normal dimorphic plumage in dimorphic species. In cockatiels, juveniles of either sex may have either yellow or grey face coloring, and this face color does not change at maturity. The loss of other juvenile features like wing spots and tail bars may be greatly delayed. As mentioned in Part 1, the development of dimorphic coloring is driven by sex hormones. The pied mutation does not appear to affect the level of sex hormones, at least not to the extent that it would cause an adult male to have female/juvenile plumage. A testosterone drop of that magnitude ought to seriously interfere with the ability to breed, but pied cocks seem to be as fertile as any other males.
I have not been able to find an explanation for why the pied mutation disrupts the normal dimorphism. But pied interferes with the normal migration of melanin-producing cells, and the plumage changes in adult males are triggered by special hormone receptors that affect the way melanin is used in the relevant area. My guess is that maybe there are specialized cells that simply fail to arrive in the right place, or fail to develop the needed hormone receptors. As mentioned earlier, Lovebirds Compendium page 499 says that some pied mutations can affect the skin cells surrounding the pigment cells, which sounds like it could play a role in th development or non-development of hormone sensitivity.
ADM pied is also the only pied mutation associated with grizzled feathers, which have an uneven mix of pigment creating a "TV static" appearance. Personally I suspect that this is related to melanoblasts that arrived erratically at this particular feather follicle. Grizzle is discussed in more detail later.
c. Progressive pied aka mottled pied
In this mutation, the number of pied feathers increases with every molt as old feathers with melanin coloring are replaced by new feathers without melanin. The pattern is random. Lovebirds Compendium page 523 indicates that this is a type of avian vitiligo caused by the interaction of several genes, and says that the pied markings are caused by the destruction of melanocytes in the feather follicle and some other tissues. It's apparently an auto-immune problem, where antibodies in the bloodstream attack the body's own pigment cells and degrade them. Progressive pied doesn't fit the standard definition in which a true mutation involves a single gene whose inheritance follows a predictable pattern. However Ornitho-Genetics lists it as an "autosomal multifactorial" mutation.
d. The pied mutation and eye color
According to Mutavi, there are two sources of melanin in the eyes. The melanin in the front of the eye has its origin in the neural crest, just like the rest of the body. But the melanin in the back of the eye has its origin in the optic cup. Pied affects the melanin that originates in the neural crest, but not the melanin that originates in the optic cup. So it doesn't usually affect the melanin in the eyes enough for us to notice the difference. But it must be possible for it to cause some reduction in the eye melanin, and in some cases we might be able to see some redness in the eye, particularly if the melanin from the optic cup happened to be sparse in a particular bird. Unexplained eye colors have been observed in lutinos (Susanne Russo; also see the Pigment in the eyes section of this article), and these colors have been observed in a more subtle form in pied birds. The lutino mutation affects all the melanin in the body, so the melanin is greatly reduced in lutino eyes, making it easier to observe any other pigments that might be present. It seems likely that the observation of odd colors in pied eyes is due to a partial reduction of melanin in the eye.
*******************
(Back to Top)
2. Opaline
Alternate common names: pearl (in cockatiels), rose (in
Bourke's parrot), red (in Eastern rosella)
Internationally agreed name: opaline
Inheritance mode: sex-linked recessive
This mutation manifests very differently in different species, to the point that I have trouble identifying it in most species. Yet it is all believed to be the result of a mutation in a single locus. It is not known whether the same allele is involved in all species, or if there are different alleles in different species that account for the variations in how the gene is expressed. When I look for information on the opaline mutation in parrot-related sources, I find a 'laundry list' of its identifying features (Martin-Parrots):
-
Melanin is redistributed, with reduced foreground melanin in most species and reduction of background melanin in some species. A few species have increased background melanin in limited areas.
-
Melanin is lost from the central areas of affected feathers, leaving a border of melanin around the edge of the feathers that varies in width from species to species and from individual to individual. This border may be wide (with an almost-normal feather appearance), or narrow (creating a very noticeable pattern).
-
Psittacin pigment is increased. The distribution of red psittacin may be greatly increased throughout the plumage (in species that already have this pigment) and yellow psittacin is also enhanced.
-
Inheritance is sex-linked.
-
Males retain the wing stripe in adulthood (in Australian parrots).
- In species where the down of chicks normally contains melanin, the melanin is lost and the down becomes thicker.
That's a very disparate set of characteristics without an obvious link between them, and what I don't find is an explanation for why or how the opaline mutation does any of these things. But the research on normal avian coloration provides a possible explanation, as described in the next section.
*******************
(Back to Top)
a.
Is it ancestral coloring?
In many species, the gradations of melanin coloring caused by the opaline
mutation are subtle, to my eyes
at least. But in pearl (opaline) cockatiels the pattern is very striking,
and it's also a lot more complex than the general description in the
previous section. Most parrot mutations display an obvious malfunction in
the color production process, but pearl looks more like a different kind of
normal. It would be wonderful
camouflage for a ground-feeding bird, and many bird species throughout the
world use similar patterning on their feathers. The markings also look much too
organized to be caused solely by an accidental change in a single
gene, without the assistance of a set of other genes to help create the pattern. I
wonder whether pearl (opaline) is a back
mutation that has restored a form of ancestral coloring.
The
Hill-McGraw volumes on bird coloration indicate that it very well could
be. Scientific research indicates that once different color patterns have been
established in an evolutionary lineage, it's not unusual to switch back
and forth between them. As a lineage evolves over time, it can go from
having dimorphic coloring to nondimorphic coloring and back again; from
having dark plumage to light plumage and back again; or from color patterns to plainer feathers and back again. Chapter 8 of
Bird Coloration Vol. 2 by Badyaev explains that the feather follicle
functions in a modular fashion, and:
"despite an amazing diversity of avian
color patterns, there is also an equally remarkable convergence of
complex pigmentation patterns among phylogenetically distinct species. Apparently, once formed, these patterns can be
preserved without being expressed for millions of generations,
suggesting that their highly modular (i.e., context-independent) genetic
and developmental organization allow these patterns to appear intact in
distinct lineages. Similarly, complex sex-specific color patterns can
lay dormant in the opposite sex and appear under hormonal treatment or
when selection against such expression ceases."
The scientific jargon in this chapter is somewhat over my head
so I don't fully understand the details. But the
important point is that a principle has been identified
by someone who DOES understand it. An internet search for feather
module badyaev will turn up a portion of the chapter on Google Books for
anyone who would like to take a look at the full discussion.
Other chapters in Hill & McGraw assert that patterned markings within a feather are always caused by melanin production switching on and off during feather growth, so I have to conclude that this is how pearl markings are produced on cockatiel feathers. The mechanism behind the on/off switch was unknown at the time these books were published, but a 2013 paper by Lin et al found that "complex patterns are not encoded directly in DNA, but result from interaction between a DNA-encoded cellular property and physical-chemical principles". They proposed several principles behind the formation of complex pigment patterns in feathers. The details aren't particularly relevant to this discussion.
Inaba & Chuong (simplified article at USC) also worked in this area, studying melanocyte progenitor cells and their effect on micro-patterns and macro-patterns in the feathers. There's also another paper on macro-patterns by Inaba et al. This is an active research area, and as new papers become available it might be possible to find them here.
There are current members of the cockatoo family who have patterned markings on their feathers, such as the red-tailed black cockatoo, the gang-gang cockatoo, and the normal hen/juvenile markings of cockatiels themselves, suggesting a common cockatoo ancestor with patterned plumage. The poorly-understood "spot gene" trait in cockatiels (aka thrush chest) appears to be another genetic pattern whose origin and relationship to pearl is unclear.
If the opaline mutation really is a reversion to ancestral plumage, then presumably the common features observed in all species that have this mutation are a legacy from a common ancestor. The differences in the way this mutation expresses itself in different species may be the result of the gene not mutating all the way back to the full original coloring in some species - in other words, the current mutation gene is an allele of the original gene in some species, and it only partly expresses the original pattern. Or differences in the subsequent evolutionary history of some species may affect the way this gene is expressed in the modern species, and some additional re-adaptation/selective breeding might be needed to fully express the original pattern.
This might not be as difficult as it looks. When the master gene for the trait was nonfunctional, in many cases there would have been no selection either for or against the modifier genes involved in the trait. Without selection for these traits, the genes could mutate freely, and it's expected that the modifiers would be "broken" in many individuals. But without selection against these traits, it's unlikely that ALL of the original wildtype alleles would have been eliminated from the population, and there could very well be some intact copies floating around. All it would take is some luck to find them.
*******************
(Back to Top)
b. Digression - are green cockatoos possible?
 |
Scientists studying the evolution of avian coloration frequently construct phylogenies (a sort of family tree) showing how a certain characteristic is distributed among different species and families of birds. As an example, see Mayr which has a series of phylogenies showing the distribution of certain types of bone structure among the parrots. These phylogenies help us reconstruct the probable characteristics of a common ancestor. When many species from the older branches of the family have trait A, and a few species from the newer branches have trait B, it is highly probable that the common ancestor had trait A, because that is much more likely than having many species evolve this trait separately.
J. Boyd has posted a simple parrot phylogeny that can be clicked on to produce a more detailed version. It shows that the split between the cockatoos and the "true" parrots occurred very early in the evolution of the parrot family. This tells us nothing about whether the common ancestor had green coloring and the cockatoos lost it at the time of the split, or if the common ancestor did not have green coloring and the other parrots acquired it at the time of the split. But fortunately there is an older branch of the family that gives us a very important clue. The Strigopidae (New Zealand parrots) branched off from the other parrots before the split between the cockatoos and the true parrots, and the Strigopidae do have green coloring.
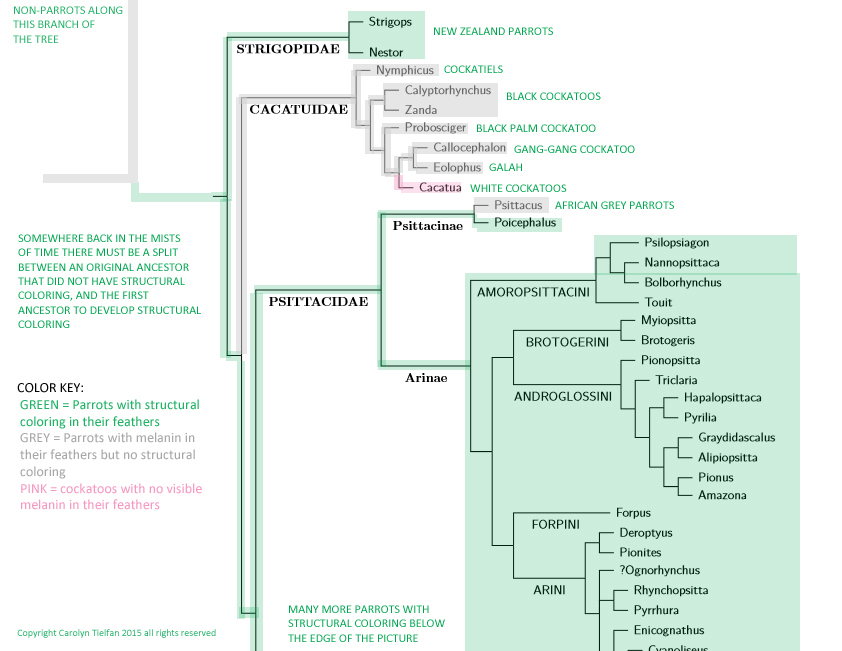 |
| My phylogeny of the most likely line of descent for structural coloring |
The normal grey coloring in cockatiels (and presumably the other cockatoos that are normally grey) might or might not be produced by the same gene that produces the grey mutation in some parrot species that are normally green. The grey mutation (discussed in a subsequent section) is dominant to the normal green coloring in these species. If it's the same gene, it's reasonable to assume that in grey cockatoos it would have to mutate into the recessive allele that produces structural coloring. Something else might be required for the black cockatoos (who apparently have their melanin on the outer layer of the feather, which would interfere with the structural color) and the white cockatoos (who don't have melanin in their feathers).
As mentioned in the previous section, complex ancestral coloration traits can be passed along intact for millions of years without being expressed, until the right set of circumstances makes them emerge again. The discussion in Hill & McGraw does not specifically mention structural coloring, but the general principle on reversion to ancestral coloring is so broad that there's no reason to think it would not apply to structural coloring. So it does seem possible that the cockatoo family might carry a package of genes related to structural coloration, just waiting for the right back mutation(s) to bring it out.
*******************
(Back to Top)
These mutations cause subtle changes in the internal structure of the feather, which alters the way light reflects from these feathers, which in turn changes our perception of the color. There are several of these mutations involving different loci. They occur only in species that normally have the ability to produce green, blue and/or purple structural coloring.
1. Dark factor
Alternate common names: dark green or jade for single factor, olive for
double factor. In birds that also have the blue mutation, alternate names
include cobalt and mauve.
Internationally agreed name: dark factor
Inheritance mode: autosomal co-dominant to the wildtype gene
This mutation reduces the thickness/development of the cloudy layer in the feather. As a single factor it reduces the thickness of the cloudy layer by about half, and with the double factor there is very little cloudy layer. As the name suggests, it makes the green (or blue) color look like a darker shade. A double factor bird has a darker color tone than a single factor bird. There appear to be several "modifier" genes which create additional variations in individual appearance. This mutation is linked to the blue gene, meaning that they both occur in different loci on the same chromosome, with an estimated crossover rate of 14% (see the Crossovers article for more information).
*******************
(Back to Top)
2. Greyy
This mutation eliminates the cloudy layer completely so that the structural
coloring disappears. If the feathers are not suffused with yellow psittacin
the resulting color will look grey instead of blue. If the feathers
ARE suffused with yellow psittacin, the coloring looks similar to the DF
dark factor aka olive, and is called greygreen because it looks greenish.
This gene is probably responsible for the normal wildtype coloring in some species like the
cockatiel and African grey parrot. As mentioned earlier, melanin coloring must have developed before
structural coloring, since structural color requires melanin in order to
function. It's likely that the dominant grey gene controlled the normal wildtype coloration
for some long-ago ancestor of the parrots. Green structural coloring must
have started out as a recessive mutation of this gene that provided such a
great survival advantage that it became the normal wildtype coloring for
most parrots. But there's still an occasional back mutation to the
older dominant gene, and since this coloring has persisted naturally in some
species it must still confer a survival advantage in some settings.
There is also a recessive grey/greygreen mutation in a couple of parrot
species. I can't find much information on it, but presumably it creates a
visual effect similar to the dominant grey mutation using a mechanism that
is at least somewhat different.
Alternate common names: greygreen
Internationally agreed name: dominant grey
Inheritance mode: autosomal dominant
*******************
(Back to Top)
3. Violet
Alternate common names: ??
Internationally agreed name: violet
Inheritance mode: autosomal co-dominant
This mutation alters the shape of the cloudy layer and the feather barb, creating a purplish effect in a bird that also has the blue mutation. The effect on color is more pronounced in a double factor bird than a single factor bird.
4. Other
structural mutations
There are a number of other structural mutations in parrots including the
following: *******************
(Back to Top)
Genetic traits are not always inheritable. Some characteristics are polygenetic, meaning that they are caused by the interaction of several genes working together. These traits are theoretically inheritable but the offspring has to get the right combination of genes from the parent, and the more genes that are involved in creating the effect, the more unpredictable the results will be.
There are also some interesting genetic anomalies that result in an individual with DNA differences in some parts of the body. But the sperm/ova produced by this individual are genetically normal, having only one type of DNA, so this "mixed DNA" can not be transmitted to the offspring.
Grizzle could be defined as a large number of small patches of two contrasting colors mixed together in a random pattern on the same feather. The effect is sort of like static on an old TV screen, or it may be described as a salt and pepper effect.
There are some bird species that have a grizzle mutation, for example the zebra finch and pigeons. In parrots it seems to be more of a special effect that occurs in some individuals for unknown reasons and can not be transmitted to offspring predictably, if at all. Martin-Parrots page 258 reports that it is associated with ADM recessive pied and not with other pied mutations. But I have not seen a reason given for why ADM pied would have this effect. Since pied interferes with the migration of melanin-producing cells, my guess would be that grizzling could occur if an area was only partly populated with melanocytes, resulting in uneven partial coloring of the feather. As reported earlier, it's believed that ADM pied has a large number of modifier genes that influence its expression, which could be the reason that grizzle isn't inheritable. With so many genes involved it's hard for the offspring to get the right combination.
Grizzling has sometimes been observed in non-pied cockatiels (and probably other species as well). There are many genes and procedures involved in the production and deposit of melanin, and anything that makes the process "stutter" could result in grizzling. Recently I've heard some statements in the cockatiel community saying that grizzle is caused by an interaction of pied and pearl. This is said to be the observation of experienced breeders, but I have doubts about the idea. I can think of a way that pearl could produce grizzle all by itself - the pearl markings are presumably caused by melanin production switching on and off during feather growth, and if the on/off switch was faulty it could produce "dribbles" of color, creating a blurred effect instead of the usual clear lines. But I can't think of any reason why the combination of pied and pearl would have this effect. It seems more likely that any grizzling in pearl pied cockatiels was caused by the pied, and pearl didn't play a role in it.
*******************
(Back to Top)
The classic halfsider is a bird with "split down the middle" coloring, where the left side of the body has a different color scheme than the right side of the body. There are other variations on this theme, such as quartersiders or smaller patches of different color with a different distribution. In parrots this coloring usually (but not always) involves blue/white coloring in part of the body and green/yellow coloring in the rest, so the blue locus is obviously involved in these cases.
The tricolor cockatiel has two different shades of grey (or grey-brown) on the body. This coloring obviously does not involve the blue locus, but otherwise it's not clear how closely related these phenomena are. Both tricolor and halfsider coloring can not be inherited by the offspring of the affected bird.
The two primary explanations for this coloring are chimerism (where fraternal twins with different coloring have merged into a single individual), and genetic mosaicism, where a DNA difference has occurred in some of the body cells of a single individual. Mosaicism is probably the best explanation for most of these cases; it's much more common than chimerism, and is more likely than chimerism to manifest with this type of pattern. I have a separate article on tricolor and halfsiders that goes into much more detail on the subject, including the possible involvement of gynandromorphy in some cases.
*******************
(Back to Top)
Almost any combination of different mutations can occur in the same bird, and it would be impossible to list all the possible variations. This section will list a small number of combinations that frequently cause confusion or that have an unexpected effect.
1. Albino (blue lutino) and creamino
The albino mutation in mammals is caused by a single gene, but this is not the case with parrots. Mammals have a single pigment in the skin, melanin, so one gene is enough to get rid of it. But parrots have two pigments - melanin and psittacin - which are controlled by different genes, so it takes the combination of two mutations to produce an all-white individual with red eyes. The blue mutation (aka whiteface) gets rid of the yellow/red psittacin coloring, while the lutino mutation gets rid of the melanin. The appearance of structural coloration disappears along with the melanin. The technically correct description of the resulting bird is a blue lutino, but it doesn't make any sense to the general public to say that a white bird is blue. So the familiar term albino is used instead. This is a good description of the bird's phenotype (physical appearance). But it doesn't convey any meaningful information about the bird's genetic makeup, which often causes confusion for people who want to know how to breed one.
"Creamino" is often used to describe a bird that combines the lutino mutation with one of the parblue mutations. Since the psittacin coloring isn't completely eliminated, a bird with yellow psittacin in the feathers has a creamy off-white appearance instead of the stark white coloring of an albino.
*******************
(Back to Top)
The lutino mutation eliminates essentially all of the visible melanin in a bird, but some of the melanin coloring comes back when cinnamon and lutino are combined. This cinnamon wash is usually pale (but can be darker) and is usually most noticeable on the primary feathers and tail. The wash is darker in adults than in juveniles, so adult hormones might play a role.
The lutino mutation severely disrupts the transportation of three enzymes, making it impossible to produce normal melanin. So very little melanin is produced, and it's small and deformed. But the cinnamon mutation eliminates one of the enzymes that needs to be transported, which apparently makes it possible to transport more of the remaining two enzymes. So more melanin is produced and deposited in the feather. This melanin is also highly abnormal, being brown colored with granules 2.5 times the normal size, and enough of it makes it into the feather that some of it becomes visible.
In budgies, the combination of cinnamon and lutino creates a pale brown "lacy" pattern on the wings in place of the usual bold black markings. So cinnamon lutino is called lacewing in budgies and sometimes in other species too. But this is rather confusing since the other species don't have similar markings and there isn't a visible "lace" effect.
*******************
(Back to Top)
In some species, reducing the melanin with either lutino or opaline increases the amount of red/orange/pink coloring. There are additional effects when opaline and lutino are combined. The word rubino is derived from rubrum, the Latin word for red.
The word rubino is used to describe this combination in species including Bourke's parakeet, rosellas, and red-rumped parrots.
4. Cinnamon fallow
The cinnamon mutation prevents melanin from completing the final transition
to black, so the color of the melanin remains brown. The bronze fallow
mutation reduces black pigment to brown in a different way, by reducing the
level of tyrosinase activity. When these two mutations are combined in
cockatiels (and also in the red-fronted kakariki), most of the melanin coloring disappears and you get a bird that
looks like a lutino. It has been reported that a color wash is not usually
visible in cockatiels, and that when a color wash is present it might look like a
different tint (apricot pinkish) in bright sunlight than the color
wash on a cinnamon lutino (which is more beige). I don't know whether this combination has the
same effect in other species.
*******************
CLICK HERE TO RETURN TO PART ONE
Copyright 2015-2021 Carolyn Tielfan all rights reserved
(Back to Top)