
Breeding & Genetics
Part One - Normal Coloration
Note: this article focuses primarily on psittacines (parrots). Some of the information is applicable to other bird and animal species.
Article sections:
PART ONE - NORMAL COLORATION
Basic elements of bird coloration
1. Feather layers
2. Melanin
3. Red/yellow pigments
a. UV markings and fluorescence
4. Structural color
5. Pigment in the eyes
a. Strange eye colors in lutino cockatiels
b. Unexplained red eyes
Color production and deposition
1. Melanin production
2. Carotenoid pigments
3. Psittacin production
4. Structural color
PART TWO - COLOR MUTATIONS IN PARROTS
Melanin-altering
mutations Psittacin-altering mutations
Distribution-altering
mutations Structure-altering
mutations
Non-inheritable genetic variations Combination
mutations
Basic elements of bird coloration Bird coloration is a lot more complicated
than mammal coloration. Mammals have one pigment, melanin, in their skin and
hair that produces the range of colors we see in these species. Birds have
melanin too, but many bird species also have a separate red/yellow pigment and many
have structural color. This is why birds can be so much more colorful than
mammals - they've got a lot more to work with.
Bird Coloration by Geoffrey Hill is an excellent, easy to read book on
the subject.
Bird Coloration Volumes 1 & 2 by Hill & McGraw are an excellent source
that is NOT easy to read, but delves into the science in much greater
detail.
There are three known types of melanin pigment. Eumelanin is
the type that parrots are known to have and it comes in two types, black and
brown.
Pheomelanin (which has alternate spellings including phaeomelanin)
has a reddish tint and produces rusty red and tawny yellow colors.
There are many bird species who have pheomelanin pigment in their
feathers, but the electron microscope studies conducted by
Mutavi have not
located this pigment in any of the parrot feathers they have studied. A more
recent study by
Oliveira Neves et al also found no pheomelanin using Raman spectroscopy
to examine the pigments in 26 species from the three main parrot lineages.
The study said "these birds seem to have evolved a capacity to avoid functional redundancy between pigments,
likely by regulating follicular gene expression". It's fairly safe to assume that parrots don't have
pheomelanin at all.
The third type, neuromelanin, is found only in the brains of
primates, especially humans, and does not play a role in coloration.
Melanin can occur in either the inner core of the feather or the outer
layer. When it's in the outer layer you see intense black coloring,
like the barring on budgie feathers. When it's in the inner core, we
see the melanin through the layer of feather structure on top of it, which
makes it look different to our eyes. Ordinary feather structure has a white
appearance, and black melanin looks grey when we see it
filtered through the white layer above it (unless special structures in the
feather act to make it look like a different color). The density of the melanin
deposited in the feather will affect the intensity of the color we see, with
more melanin producing darker colors and less melanin producing pale
colors.
Most bird species including canaries and finches get the red/yellow coloring
in their feathers from carotenoid pigment in the diet, especially the
xanthophyll carotenoids, although more familiar types like beta carotene
also play a role. Parrots don't use dietary carotenoids for feather
coloring. Instead they have a unique pigment called
psittacofulvin (aka psittacin) that they manufacture in the body. There are a few other bird families that make a special yellow/red
pigment, for example penguins have their own unique pigment.
"Psittacofulvin" is really the best word for this pigment, since it can't be
mistaken for anything else, while the word "psittacin" is much too similar
to the word "psittacine". But "psittacin" is the word that's most
commonly used (possibly because it's easier to type) so that is the word
that will be used in the rest of the article.
Psittacin pigment is found only in the outer layer of the feather.
For parrot species that have red/yellow pigment in their eyes and/or beaks,
there are indications that the pigment might not be psittacin. The blue
mutation in parrots removes the psittacin pigment from the feathers but
apparently does not affect beaks and eyes. The male blue-mutation
eclectus parrot still has an orange/yellow beak, and the blue mutation
princess parrot still has a pink beak and orange eyes. However it's possible
that the blue mutation only affects the deposition of psittacin in the
feathers, but does not interfere with it in other parts of the body.
UV markings and fluorescent color. Many bird species (including most if not all parrots) can see
light waves in the near-UVA
part of the spectrum, which is a part of the spectrum that humans cannot see (Hausmann
et al). Many birds also have markings that can only be seen under UV light, and some species have markings which are not just
UV-sensitive but are actually fluorescent. Some species have
fluorescent markings on their beaks (Atlas
Obscura).
A doctoral dissertation by
Pohland has a chapter on fluorescent feathers starting on page 79.
Most sources attribute these UV-sensitive markings to pigment alone, but Hausmann
et al states that "many UV-reflective plumage signals are created, in
part at least, by the microstructure of the feathers rather than by
pigmentation".
UV markings and fluorescence are usually found on body parts that are used in
sexual display. The nature of UV-sensitive pigments is poorly understood
(Pohland
page 81). But
Arnold et al say the biochemical pathway for creating fluorescent
pigments is complex and costly enough that fluorescent feathers may be an
honest signal indicating individual quality.
Many parrot species have fluorescent yellow
feathers, but fluorescent red feathers apparently do not occur in parrots
even though psittacin pigment is used in both red and yellow feathers (McGraw
& Nogare). Assuming that fluorescence in parrots is created with
pigment alone, then presumably the fluorescent yellow feathers use a form of psittacin that is somewhat different from the non-fluorescent yellow
feathers.
Mutavi hypothesizes about the
role of enzymes in producing these different types of yellow feathers, but the accuracy
of this idea is
unknown.
Pages 94-100 of
Pohland lists the species in which fluorescent plumage has been detected. Cockatiels are on
page 96.
Structural color The colors in the visual spectrum are
traditionally described as red, yellow, orange, green, blue, indigo and
violet. Red has the longest wavelength and violet has the shortest.
Notice that in birds, the longer-wave colors (red, yellow, orange) are
produced using pigment and the shorter-wave colors (green, blue, purple) are
created with an optical illusion. That's true of non-iridescent color,
anyway. Iridescent color is always structural, even the reds.
The structural layer (aka spongy layer or cloudy layer)
is the middle layer of the feather.
Many bird species have special pigments in the eye that aren't found in the feathers
or anywhere else in the body. Many pigments have other functions in
addition to coloration, and the pigments in the eye may help protect the eye
and enhance vision. The eye color of some parrots makes it
seem likely that at least some species have additional pigments in
the eye, but the pigmentation of parrot eyes hasn't been studied so I have
no information on it.
It's possible for the eyes to have structural color as well, and blue eyes
in humans are structural color.
Strange colors in the eyes of lutino cockatiels
Some strange, unexplained color effects have
been observed in the eyes of lutino cockatiels, and to a lesser degree in
other cockatiel mutations (Susanne
Russo). The odd eye colors include white and pale shades of blue, grey, green,
and brown. Sometimes there isn't any unexpected color. So there seems to be something going on in the eye that isn't
happening in the rest of the body. It's hypothesized that specific eye
colors are related to the other mutation genes that the cockatiel has, but I
don't see how these genes could cause these specific effects.
My personal
guess is that there could be remnants of other pigments, structural color,
or some other factor in a
cockatiel's eye that's related to now-superseded ancestral eye coloring.
Brown eyes are the normal coloring nowadays, so it would be hard to see
these vestigial colors when the normal melanin is present. But these colors
could become visible when the melanin in the eye is reduced. And when the
color becomes visible, there could be a lot of individual variation - or no
color at all -
because nature is no longer selecting for anything specific.
My best hypothesis at present is related to a feature found in a few
parrots like budgies and Indian ringnecks: the white iris ring that develops
in adults. Cockatiels don't have an eye ring now, but it's possible
that their ancestors did, and many individuals might still carry remnants of it. In species that normally have a white eye ring, it’s a trait that develops at maturity. This could explain why we only see the odd colors in adult cockatiels, not in chicks.
According to a
Terry Martin article on fallow mutations, the white iris ring is caused by lipid deposits
in the eye (I haven't been able to confirm this). Martin says furthermore
that in budgies and a couple of other species, one fallow mutation retains
the eye ring but the trait is lost in the other fallow mutations. So it
looks like the loss of the eye ring may sometimes be related to changes in
melanin. I haven't found a direct explanation of a relationship between melanin and lipids, but several scientific papers seem to indicate that natural melanin is a polymer made of melanin granules, protein, and lipids bound together. So it’s plausible that
the same mutation could affect both melanin and the eye ring because of its
effect on the lipid component.
There is a type of albinism in rabbits
called the Vienna gene, which produces individuals with white fur and blue
eyes. It doesn't appear to have any relevance to the odd eye colors
observed in lutino cockatiels (Deb's
Rabbit Tree).
Unexplained red eyes. Sometimes an
individual bird has red eyes for no known reason. According to
Mutavi, there are two sources of melanin in the eyes. The melanin in the
front of the eye has its origin in the neural crest (the embryonic precursor
of the spinal cord), which is also where the melanin in the skin and
feathers originated. But the melanin in the back of the eye has its origin
in the optic cup. A mutation that affects all the melanin in the body (like
lutino) will affect all the melanin in the eye. A mutation that
affects only the melanin that originated from the neural crest (like pied)
is expected to affect some of the melanin in the eye. But it does not
affect the melanin that originated in the optic cup, so there's usually not
a noticeable difference in eye color. It seems possible that an unidentified mutation could affect the melanin
originating in the optic cup without affecting the melanin in the rest of the body,
resulting in a red-eyed bird that we don't understand.
1. Sex-linked lutino
a. Parino mutations
(lutino alleles) - lime/pallid/platinum
2. Cinnamon
3.
Fallow mutations (including recessive silver and NSL ino)
4.
Dilute mutations (including emerald)
5. Dominant edged (including dominant silver)
6. Faded/Isabel
7. Melanistic
1. Blue (whiteface)
a. Parblue mutations
2. Orangeface/tangerine/dominant yellow cheek
3. Sex-linked yellow
cheek and goldcheek
4. Red-fronted and
red suffused
1. Pied
a. Dominant
b. Recessive
c. Progressive
d. The pied mutation and eye color
2. Opaline/pearl
a. Is it ancestral
coloring?
b. Digression - are green
cockatoos possible?
1. Dark factor/olive
2. Grey/greygreen
3. Violet
4.
Other structural mutations
1. Grizzle
2. Halfsiders and tricolor
1.
Albino (blue lutino), creamino (parblue lutino)
2. Lacewing (cinnamon lutino)
3. Rubino (opaline lutino)
4. Cinnamon fallow
 The
elements of coloration aren't distributed equally throughout the feather;
instead they occur in three different layers, called the medulla (inner
core), the cloudy layer, and the cortex (outer layer). Click on the
thumbnail at left to see cross-section diagrams of typical feathers from
Terry Martins' parrot mutations book.
The
elements of coloration aren't distributed equally throughout the feather;
instead they occur in three different layers, called the medulla (inner
core), the cloudy layer, and the cortex (outer layer). Click on the
thumbnail at left to see cross-section diagrams of typical feathers from
Terry Martins' parrot mutations book.
Taysom et al report that psittacin coloring appears to be
physiologically less costly than carotenoid coloring, and also that
psittacin coloring doesn't seem to be a major factor in sexual selection. In
general, the costliest coloration plays the biggest role in sexual selection
because this is the best indicator of the individual's condition. With
parrots, structural color apparently plays a bigger role in sexual selection
than melanin or psittacin pigment does. But the study did not include the
cockatoo family, which does not have structural color. It also did not
discuss fluorescence, which may play a role in sexual selection.
There are special types of feather structure that can create the appearance
of other colors and/or iridescence.
Blue, green and purple are usually produced by structural coloring which is
basically an optical illusion. There are no known blue or purple pigments in birds,
and the only known green pigment is in turacos.
It has been observed that the blue structural color in scarlet macaw
wing feathers does not use the standard technique: "Unlike many
other birds, the blue coloration of scarlet macaw feathers stems from
feather barbs rather than barbules... The adoption of this rod-volume
fraction by feather barbs to attain the highest brightness of the blue
coloration is a nontrivial optimization, apparently a result of natural
evolution." (Yin
et al). I have no information on whether other structural
color in parrots is achieved the same way, for example the green body
feathers of an Amazon parrot.
*******************
(Back to Top)
Color production and deposition
Most color mutations involve a disruption to the normal coloration process. Something new has not been added to the bird's coloring; instead, some part of the normal coloration process has broken down so that normal coloring can not be produced. The appearance of the mutation coloring will depend on the nature of the malfunction. It is helpful to understand how normal coloring is produced in order to understand the effect that mutations have.
Feathers grow from a structure in the skin called the feather follicle. The feather follicle is shaped like a ring or collar and is equipped with a rich blood supply, nerve endings, muscles to control the position of the feather, and specialized cells related to feather growth. The cylindrical shaft of a new feather grows from the center of the feather follicle. This article will not discuss every detail of feather growth, just the aspects related to coloration.
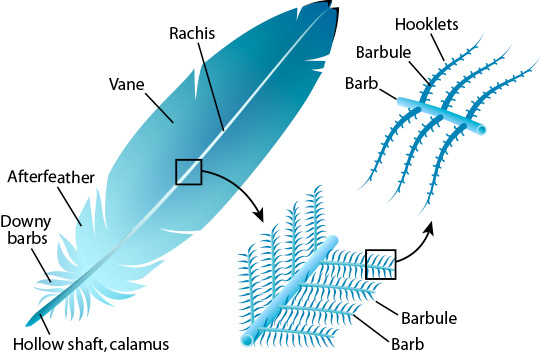 Feathers
are not made of living cells; they are mostly composed of keratin, the same
inert material that hair and fingernails are made of. Pigment is
deposited in a new feather while it is actively growing, and any special
features needed for structural coloration are also created at this time.
Pigment is deposited primarily in the barbs and barbules of the feather vane. Once the
feather has finished growing, there isn't any transportation of material
into or out of the feather. Additional pigment can not be added to a
fully grown feather, and the existing pigment can not be "brightened up" by
changes in health or diet. However the appearance of the feather can be
improved by better grooming (including preening and bathing), or made worse
by dirt, damage or natural fading.
Feathers
are not made of living cells; they are mostly composed of keratin, the same
inert material that hair and fingernails are made of. Pigment is
deposited in a new feather while it is actively growing, and any special
features needed for structural coloration are also created at this time.
Pigment is deposited primarily in the barbs and barbules of the feather vane. Once the
feather has finished growing, there isn't any transportation of material
into or out of the feather. Additional pigment can not be added to a
fully grown feather, and the existing pigment can not be "brightened up" by
changes in health or diet. However the appearance of the feather can be
improved by better grooming (including preening and bathing), or made worse
by dirt, damage or natural fading.
It was surprisingly difficult to find information on melanin production that was clear, concise and complete. The sources for the description here include Yu et al, Genetics and Evolution of Domestic Fowl, Palaeobiology Research Group, Lin et al, Cichorek et al, Slominski et al, Mutavi (Onsman), Mutavi (Van den Abeele), Cock and Cohen, Eberhard, and Wikipedia. The description here is focused on eumelanin, since that is the form of melanin used by parrots. The production of pheomelanin is similar, except that the amino acid cysteine is incorporated at a specific stage of the process.
The potential to produce melanin begins in the embryo when nonpigmented melanoblasts (precursors to melanin-producing cells) arise in the neural crest (the area that will become the spinal cord) at about the level of the shoulders and migrate to the skin. At first they are evenly distributed, but then they become localized in the "feather primordia", the specific areas where feathers will grow. The melanoblasts persist and multiply here and disappear from non-feather-related areas of the skin. The melanoblasts develop into melanocytes, which are mature cells capable of melanin production.
The melanocytes congregate beneath the feather germs
which lie at the base of the feather follicles, adjacent to another type of
cell called keratinocytes. Initially the melanocytes are randomly arranged in the
proximal feather germ, but they gradually become aligned in parallel
rows as barb ridges form. They connect to the growing feather and deliver melanin for a time,
then the connection is broken and the melanocyte migrates and dies. The number of melanocytes in each barb ridge varies from one to four.
Therefore, unlike humans whose melanocytes are long-lived, birds require a
reservoir of melanoblasts (stem cells) which can develop into melanocytes in
the future to
be used with new feathers. This reservoir is probably located at the base
of the feather follicle, although some suggest that new melanocytes may
also come from extra-follicular melanoblasts.
Normal melanin is produced by
oxidation of the amino acid tyrosine. The enzyme tyrosinase
plays a leading role in the process. A variety of
other chemical compounds are involved as well; the ones that are known to be directly
relevant to mutation coloring in parrots include the MATP protein and the TRP1 and TRP2
enzymes/proteins.
The granules of melanin pigment are created through the development of melanosomes (membrane-encased organelles
that arise in the melanocytes during the melanin production process). The
number of words beginning with "melano" can be rather confusing; to clarify,
the melanocyte ("melanin cell") is the name for the entire cell where melanin is produced.
The melanosome ("melanin body") essentially develops into a package full of
melanin pigment that will be deposited in the feather. Melanocytes are
branched structures, consisting of a central cell body with a large number
of
branches or tubules called dendrites. The melanosomes (packages of melanin
pigment) begin their
development in the middle of
the melanocyte cell, migrating to the outer edge of the cell through the
dendrites as development proceeds. In species that produce both eumelanin
and pheomelanin, the melanocytes may produce eumelanosomes or phaeomelanosomes at
different times, switching from one to the other. There are four stages in
the physical production of melanin. In Stage 1, the organization of a
colorless protein matrix begins. At this stage the organelle is called a premelanosome
and it is a small, round vesicle with an amorphous matrix. In Stage 2,
it has developed into a melanosome with a changed shape and an
organized, structured matrix. Tyrosinase has appeared but pigment
synthesis has not yet begun. Pigment production begins in Stage 3 and
the pigment is deposited on the matrix. There is a high level of
tyrosinase activity at this stage. In Stage 4, the whole melanosome is filled with pigment. Tyrosinase activity
slows down and ceases.
The transport protein myosin helps
transfer the melanosome to the keratinocytes for deposition into the
developing feather. Melanosome transfer is
a cytophagic ("cell engulfing") process during which a portion of a
melanocyte dendrite is pinched off by the epidermal cell so that melanosomes
and melanocyte cytoplasm are incorporated into the keratinocyte.
Keratinocytes are the cells that produce the protein keratin, the key
structural component of feathers.
Once the pigment package has left the melanin-producing cell, it is referred
to simply as melanin not a melanosome. Here is
Wikipedia's
description of the chemical
process involved in melanin production: The first step of the biosynthetic pathway for both eumelanins and
pheomelanins is catalysed by tyrosinase:
Dopaquinone can combine with
cysteine by two pathways to benzothiazines and pheomelanins: Also, dopaquinone can be converted to
leucodopachrome and follow two more pathways to the eumelanins:
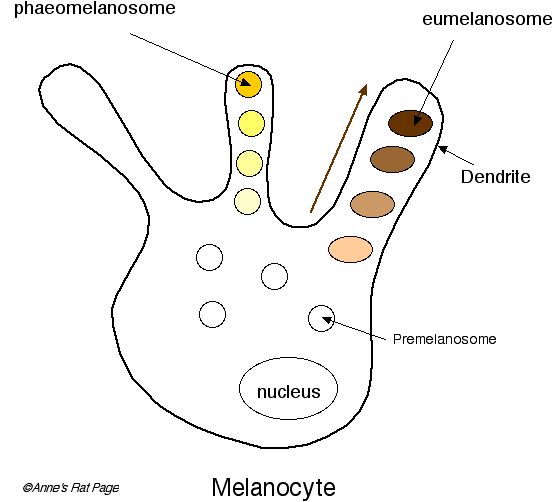
Diagram of a melanocyte from
Palaeobiology Research Group
Tyrosine →
DOPA →
dopaquinone
Dopaquinone + cysteine → 5-S-cysteinyldopa → benzothiazine
intermediate → pheomelanin
Dopaquinone + cysteine → 2-S-cysteinyldopa → benzothiazine
intermediate → pheomelanin
Dopaquinone → leucodopachrome →
dopachrome→ 5,6-dihydroxyindole-2-carboxylic acid →
quinone →
eumelanin
Dopaquinone → leucodopachrome → dopachrome → 5,6-dihydroxyindole →
quinone
→ eumelanin
The production of melanin can be finely controlled during feather growth, switching on and off in response to chemical signals. This is what allows the creation of intricate color patterns on feathers.
Carotenoid pigments*******************
(Back to Top)
This subject won't be discussed in detail since parrots don't use carotenoid pigments for coloration. Carotenoid pigments come in two basic colors - red and yellow - and related colors like orange and pink come from a mix of the red and yellow pigments or a combination with structural white. Most carotenoids in food sources are yellow although red carotenoids can be obtained from some foods. Most of these dietary carotenoids are deposited directly into the growing feather with no further modification, although some bird species have the ability to process yellow carotenoids into red within the body. The type of carotenoids used for feather coloration may vary by species, but in general the most important types are two xanthophyll carotenoids called lutein and zeaxanthin. The type and amount of carotenoids eaten by the bird has a direct effect on the intensity of feather coloring (especially in males). Carotenoid coloring plays a role in mate selection for many species because it is a strong indicator of a male's ability to find food or defend a territory with good food resources.
Unlike melanin coloring, carotenoid coloring is not finely controlled by the body. By themselves, carotenoids can be deposited into the feathers in large blocks but it is believed that they can not be used to create fine patterns. When you see a feather with an alternating pattern of black and yellow (for example), it was the black melanin that controlled the pattern not the yellow carotenoids.
Although parrots don't use carotenoid pigments to color their feathers, carotenoids have many other functions in addition to coloration and parrots do use carotenoids in other ways, and they have carotenoids circulating in the bloodstream. Probably everyone knows that beta carotene is the precursor to vitamin A. Xanthophyll carotenoids provide the yellow/orange color in egg yolks, and it's obviously not for decorative purposes since egg yolks aren't meant to be seen. It's thought that the carotenoids help protect the embryo from oxidative damage, and studies have found that hens with more carotenoids in their egg yolks have better hatch rates and better fledging success. It's likely that parrots have xanthophyll carotenoids in the eye, since these substances have a protective function in the eye and even humans have lutein and zeaxanthin in the eye.
*******************
(Back to Top)
Very little is known about psittacin. Like carotenoids, psittacin comes in two basic colors (yellow and red) and related colors are created by blending the two with each other or with white.
Unlike carotenoids, psittacin is synthesized in the body. At this point we don't know how it's done. Psittacin does not circulate in the bloodstream, so presumably it is synthesized in the feather follicle and not created somewhere else in the body and transported to the follicle (McGraw). Presumably it is NOT synthesized in the melanocytes, since the pied mutation (which causes melanocytes to be completely absent from some areas of the body) does not remove the psittacin from these areas along with the melanin.
The chemical nature of psittacin is poorly understood as well. It has been observed that 44 species of parrots from all three three major psittaciform families used the same set of five polyenal lipochromes to create red coloring in feathers (McGraw). But the chemical structure of yellow psittacin has resisted identification and isn't known at present.
It is unclear whether psittacin pigment can be used to create intricate color patterns on feathers. In general it seems that parrots do not use it in this way. However there are some parrots with simple psittacin patterns on their feathers, for example the crest feathers in the Major Mitchell's cockatoo has alternate bands of red, orange, and pink, and it doesn't look like melanin was involved at all in creating this pattern.
It appears that all parrot species use psittacin pigment in at least some of their feathers so this pigment is widespread in the parrot family. But it isn't known to occur anywhere else in nature - it is unique to parrots.
The precise location and method for psittacin production has not been identified, but it seems likely that the process has a superficial similarity to melanin production. Presumably psittacin is synthesized from a variety of materials in a multi-stage process and is then deposited in the growing feather, and this process is controlled by a number of different genes. A mutation in any one of these genes could disrupt the normal process in a way that affects the quality, quantity or distribution of psittacin.
Bonin and Homberger report that yellow psittacin reflects light somewhat differently in cockatoos than in the other parrots they tested. However, their conclusion that this might be an indicator of convergent evolution seems doubtful in the extreme. Psittacin pigment occurs in all the major parrot groups, including the kea of the New Zealand parrots (McGraw) and absolutely nowhere else in nature, which seems like a strong argument in favor of a common ancestor and against convergent evolution. Also, fluorescence is found in many parrots not just the cockatoos, but it is found only in parrots and not in any other type of bird (Hausmann et al).
There is ongoing research on psittacin and other aspects of avian coloration at the McGraw Lab at Arizona State University.
*******************
(Back to Top)
The three main types of structural color are white, iridescent and non-iridescent.
White is the default color of the feather, the color that we see when no pigments are present. It is the result of microstructures in the feather barbs scattering light of all wavelengths equally and randomly.
With the other types of structural color, the scattering of light is neither equal nor random; the feather has been "designed" by nature to produce a specific visual effect. With iridescent coloring, the appearance of the color varies depending on viewing angle. At some angles the color is brilliant and at other angles it disappears entirely, looking like dull dark ordinary feathers. At some angles it might look like an entirely different color from what is expected, for example green instead of red. With non-iridescent structural coloring, the color looks the same from all viewing angles. Both types achieve their effect by using structures within the feather to affect the way that the light hitting the feathers is reflected back to the viewer.
Iridescent coloring is produced by light reflecting from crystal-like arrays in the feather barbules, consisting of thin layers of keratin, melanin and air. I won't go into detail since parrots generally don't use iridescent coloring, but this paper at the Journal of Experimental Biology has additional information.
Non-iridescent structural coloring is produced by microstructures in the feather barbs. Structures of keratin protein interspersed with air pockets in the spongy layer of the feather allow short-wave blue and violet light to be reflected back to the viewer, while longer wavelengths are absorbed by the melanin in the feather's core. The shade of blue is determined by the shape and size of the air pockets (Smithsonian, The Scientist, Hill & McGraw). The way that the light is reflected back was originally believed to be Tyndall scattering (aka Rayleigh scattering) but is now known to be constructive interference. With constructive interference the light is not simply reflected back; some wavelengths are actually amplified. If there is yellow pigment in the outer layer of the feather, the combination of blue and yellow will look like green, and if there is red pigment in the outer layer it will look like purple. The melanin layer in the feather core is necessary to get this effect, and the structural layer can't function properly in a mutation that eliminates the melanin. A feather with no pigment in it will simply look white whether it has the structural layer or not.
The special feather structure in parrots is sometimes called Dyck texture, after researcher Jan Dyck who studied the phenomenon. Most parrot species have non-iridescent structural coloration, with African greys and all members of the cockatoo family being the major exceptions. It is probably not a coincidence that these are the same species that use powder down to groom their feathers; presumably this powdery substance would interfere with the visual effect of structural color if it was present.
Structural color can occur in other parts of the integument (the outer covering of the body) such as exposed skin on the face or legs, the beak, or the eyes.
*******************
(Back to Top)
Dimorphic coloring and other normal plumage variations
In some bird species, including many parrot species, all individuals who are old enough to have feathers look more or less alike. You can't tell males from females or juveniles from adults just by looking at them. It's called monomorphism ("one form") or monochromatic coloring when all adults have the same basic plumage pattern.
But there are also many bird species including some parrots that normally have plumage differences which indicate the sex or age of an individual. It's called sexual dimorphism ("two forms") or dichromatic coloring when adult males look different than adult females. In many dimorphic species, all juveniles look the same as females and the males acquire their distinctive plumage when they mature. But in some species, juveniles have their own distinctive plumage that is different from either parent (whether the adults are dimorphic or not). There are also species where the young go through a series of different plumages, changing from molt to molt until they reach full adult plumage after a period of several years. This is called delayed plumage maturation (DPM), and it is not a sign of sexual immaturity; subadult coloring is associated with individuals who are sexually capable. There are also species who have different plumage at different times of year, either for camouflage purposes or because special plumage during the breeding season helps with mate attraction.
In all of these cases, the plumage differences are regulated by hormones. The hormones involved vary from species to species. Since sexually dimorphic coloring is common in parrots but the other variations are not, sexual dimorphism is the only type of plumage difference that will be discussed here.
Dimorphic coloring is regulated by sex hormones, especially testosterone and estrogen. In most bird species including parrots, it appears that testosterone is the controlling factor. Female coloration is the default appearance but the elevated testosterone levels of an adult male causes a change. This is why juveniles of both sexes look the same as adult females in many species; the juvenile males don't have enough testosterone yet to trigger the development of male plumage. The precise way that sex hormones influence plumage color isn't fully understood, and may vary from one species to another. But it appears that the feather follicle (or something in it) is sensitive to the relevant hormone and responds differently to different hormone levels (Goymann & Wingfield). There are cells in many areas of the body with special hormone receptors that have a specific reaction to the appropriate hormone level (Wikipedia), and presumably these specialized hormone receptors are involved in the development of dimorphic plumage.
There are some species (particularly gallinaceous birds like waterfowl and quail) where male appearance is the default and the influence of estrogen makes the female plumage pattern appear. There are also some species where the hormone that makes the difference is not estrogen or testosterone, but some other breeding-related hormone like luteinizing hormone (LH).
Hormonal imbalances can make an individual have the plumage of the "wrong" sex in part or in full. In species where testosterone is the controlling hormone, insufficient testosterone can make a male acquire female-type plumage and excessive testosterone can make a female acquire male-type plumage. A similar principle applies to species where estrogen is the controlling hormone. The influence of hormones isn't limited to the feathers; it can affect other dimorphic color traits, for example cere color in budgies. Budgies with severe hormone imbalances have been observed to have the cere color of the opposite sex.
Eclectus parrots are an interesting case. They have the most extreme dimorphic coloring in the parrot family, with males having mostly bright green feathers and an orange/yellow beak, while females have red and blue/purple feathers with a black beak. I couldn't find any information on what causes the color difference in this species, but I would guess that it isn't testosterone or estrogen. The color difference between males and females is present when the chicks begin to grow feathers, long before they develop significant levels of breeding hormones.
*******************
(Back to Top)
CLICK HERE TO CONTINUE TO PART TWO
Copyright 2015-2021 Carolyn Tielfan all rights reserved